Dela med sig:



Vad är replikationsgaffeln?
den replikationsgaffel Det är punkten vid vilken DNA-replikation sker, det kallas också en tillväxtpunkt. Den har en Y-form, och efterföljande replikation förskjuts hårnålen av DNA-molekylen.
DNA-replikation är den cellulära processen som involverar duplicering av genetiskt material i cellen. Strukturen av DNA är en dubbel helix, och för att replikera dess innehåll måste den öppnas. Var och en av strängarna kommer att ingå i den nya DNA-kedjan, eftersom replikation är en semiconservativ process.
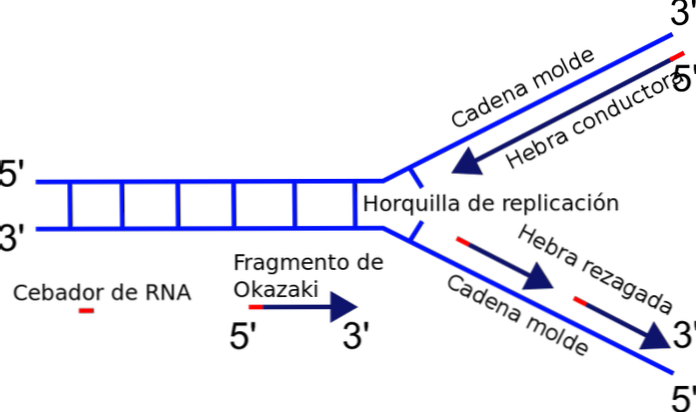
Replikeringsgaffeln bildas strax mellan korsningen mellan den nyligen separerade mallen eller mallkedjorna och duplex-DNA som ännu inte har duplicerats. När initiering av DNA-replikation kan en av trådarna enkelt dupliceras, medan den andra strängen står inför ett polaritetsproblem.
Det enzym som är ansvarigt för polymerisering av kedjan - DNA-polymeraset - syntetiserar endast DNA-strängen i 5'-3'-riktningen. Således är en sträng kontinuerlig och den andra lider en diskontinuerlig replikation, vilket genererar fragment av Okazaki.
index
- 1 Replikering av DNA och replikationsgaffel
- 1.1 Enriktad och dubbelriktad replikering
- 1.2 Enzymer involverade
- 1.3 Start av replikering och gaffelbildning
- 1.4 Förlängning och rörelse av gaffeln
- 1.5 Avslutande
- 2 DNA-replikation är semikonservativ
- 3 Polaritetsproblemet
- 3.1 Hur polymeras fungerar?
- 3.2 Produktion av Okazaki-fragment
- 4 referenser
Replikering av DNA och replikationsgaffel
DNA är molekylen som innehåller den nödvändiga genetiska informationen för alla levande organismer - med undantag för vissa virus.
Denna stora polymer sammansatt av fyra olika nukleotider (A, T, G och C) ligger i kärnan hos eukaryota, i var och en av cellerna som utgör vävnaderna i dessa varelser (utom mogna röda blodceller från däggdjur, som saknar av kärnan).
Varje gång en cell delar upp, måste DNA replikeras för att kunna upprätta en dottercell med genetiskt material.
Enriktad och dubbelriktad replikering
Replikationen kan vara enriktad eller dubbelriktad, beroende på bildningen av replikationsgaffeln vid ursprungspunkten.
Logiskt är, i fallet med replikering i en riktning, en enda gaffel bildad, medan två forkrifter bildas i tvåvägsreplikation.
Enzymer involverade
För denna process behövs en komplex enzymatisk maskin som fungerar snabbt och som kan replikera DNA på ett exakt sätt. De viktigaste enzymerna är DNA-polymeras, DNA-primas, DNA-helikas, DNA-ligas och topoisomeras.
Start av replikering och formning av gaffeln
DNA-replikation startar inte på någon slumpmässig plats i molekylen. Det finns specifika regioner i DNA som markerar början på replikationen.
I de flesta bakterier har bakteriekromosomen en enda AT-rik utgångspunkt. Denna komposition är logisk eftersom den underlättar öppningen av regionen (AT-paren är förenade med två vätebroar medan GC-paret med tre).
När DNA: n börjar öppna, bildar en Y-formad struktur: replikationsgaffeln.
Förlängning och rörelse av gaffeln
DNA-polymeras kan inte starta syntesen av dotterns kedjor från början. Du behöver en molekyl som har en 3'-ände så att polymeraset har att börja polymerisera.
Denna fria 3'-änden erbjuds av en liten molekyl av nukleotider som kallas primer eller primer. Den första verkar som ett slags krok för polymeras.
Med replikationsförloppet har replikationsgaffeln förmågan att röra sig längs DNA: n. Plattformen av replikationsgaffeln lämnar två enbands-DNA-molekyler som styr bildningen av dubbelbandsmodulmolekylerna.
Gaffeln kan avancera tack vare åtgärden av helikasenzymerna som uncoil DNA-molekylen. Detta enzym bryter vätebindningarna mellan basparen och möjliggör förskjutningen av gaffeln.
fullbordan
Replikationen avslutas när de två gafflarna är vid 180 ° C från ursprunget.
I det här fallet talar vi om som flyter processen att replikera i bakterier och det bör noteras att processen med att vrida den cirkulära molekylen involverar replikering. Topoisomeraser spelar en viktig roll vid avvecklingen av molekylen.
DNA-replikation är semikonservativ
Har du någonsin undrat hur replikation sker i DNA? Det vill säga att en annan dubbelhelix måste uppstå från dubbelhelixen, men hur händer det? I flera år var det en öppen fråga bland biologer. Det kan finnas flera permutationer: två gamla strängar tillsammans och två nya tillsammans, eller en ny tråd och en gammal för att bilda dubbelspiralen.
1957 svarades denna fråga av forskare Matthew Meselson och Franklin Stahl. Den replikationsmodell som föreslagits av författarna var semikonservativ.
Meselson och Stahl uppgav att resultatet av replikering är två dubbelsträngade DNA-molekyler. Var och en av de resulterande molekylerna består av en gammal sträng (från moderen eller den ursprungliga molekylen) och en ny syntetiserad ny sträng..
Polaritetsproblemet
Hur polymeras fungerar?
DNA-helixen är bildad av två kedjor som löper på ett antiparallellt sätt: en går i 5'-3'-riktningen och en annan 3'-5 '.
Den mest framträdande enzymreplikeringsprocessen är DNA-polymeras, som ansvarar för katalys av bindning av nya nukleotider kommer att läggas till kedjan. DNA-polymeraset kan endast förlänga kedjan i 5'-3'-riktningen. Detta faktum hindrar samtidig duplicering av kedjorna i replikationsgaffeln.
Varför? Tillsatsen av nukleotider sker vid den fria änden 3 'där en hydroxylgrupp (-OH) finns. Således kan endast en av kedjorna lätt amplifieras genom terminal tillsats av nukleotiden till 3'-änden. Detta kallas ledande eller kontinuerlig sträng.
Framställning av Okazaki-fragment
Den andra strängen kan inte långsträckta, eftersom den fria änden är 5 'och inte den 3' och varje polymeras katalyserar additionen av nukleotider till 5'-änden. Problemet löses med syntesen av multipla korta fragment (nukleotider 130 till 200), var och en i den riktning som är normal replikation 5 'till 3'.
Denna diskontinuerliga syntes av fragment slutar med föreningen av var och en av delarna, en reaktion katalyserad av DNA-ligas. För att hedra upptäckten av denna mekanism, Reiji Okazaki, kallas de små segmenten syntetiserade fragment av Okazaki.
referenser
- Alberts, B., Bray, D., Hopkin, K., Johnson, A.D., Lewis, J., Raff, M., ... & Walter, P. (2015). Väsentlig cellbiologi. Garland Science.
- Cann, I. K., & Ishino, Y. (1999). Archaeal DNA-replikation: identifiera bitarna för att lösa ett pussel. Genetics, 152(4), 1249-67.
- Cooper, G. M., & Hausman, R.E. (2004). Cellen: Molekylär tillvägagångssätt. Medicinska naklada.
- Garcia-Diaz, M. & Bebenek, K. (2007). Multipla funktioner av DNA-polymeraser. Kritiska recensioner inom växtvetenskap, 26(2), 105-122.
- Lewin, B. (2008). generna IX. Mc Graw-Hill Interamericana.
- Shcherbakova, P.V., Bebenek, K., & Kunkel, T. A. (2003). Funktioner av eukaryota DNA-polymeraser. Vetenskapens SAGE KE, 2003(8), 3.
- Steitz, T. A. (1999). DNA-polymeraser: strukturell mångfald och gemensamma mekanismer. Journal of Biological Chemistry, 274(25), 17395-17398.
- Watson, J. D. (2006). Molekylärbiologi hos genen. Ed. Panamericana Medical.
- Wu, S., Beard, W. A., Pedersen, L.G., och Wilson, S.H. (2013). Strukturell jämförelse av DNA-polymerasarkitektur föreslår en nukleotidgateway till polymerasaktiv plats. Kemiska recensioner, 114(5), 2759-74.